Insertional Mutagenesis and Autoimmunity Induced Disease Caused by Human Fetal and Retroviral Residual Toxins in Vaccines por Peter Jarzyna (Ph.D.), Ngoc V. Doan (B.S.) y Theresa A. Deisher (Ph.D.), vinculados al Sound Choice Pharmaceutical Institute. https://www.researchgate.net/publication/326378590_Insertional_mutagenesis_and_autoimmunity_induced_disease_caused_by_human_fetal_and_retroviral_residual_toxins_in_vaccines

Contenidos
- 1. Introducción al estudio
- 2. Contenido en algunas vacunas
- 3. El problema de los residuos de ADN fetal humano
- 3.1. ¿De dónde viene ese ADN?
- 3.2. Cantidades que superan los límites recomendados
- 3.3. Fragmentación: un intento de solución que pudo empeorar las cosas
- 4. Mutagénesis insercional: Cuando el ADN extraño se introduce en nuestros genes
- 4.1. ¿Cómo entra el ADN contaminante a nuestras células?
- 4.2. La conexión con el autismo y otros trastornos del neurodesarrollo
- 5. Autoinmunidad: Cuando el cuerpo se ataca a sí mismo
- 6. El papel del retrovirus HERV-K
- 7. El momento crítico: La ventana de vulnerabilidad en la infancia
- 8. El ejemplo de la terapia génica que salió mal
- 9. Recomendaciones de los autores
- 10. Referencias
1. Introducción al estudio
El estudio aborda una preocupación que rara vez se discute en los medios de comunicación ni en los prospectos de las vacunas: los residuos de ADN fetal humano que quedan como contaminantes en ciertas vacunas fabricadas con líneas celulares derivadas de fetos abortados, y los posibles daños que estos residuos podrían causar mediante dos mecanismos biológicos bien establecidos: la mutagénesis insercional y la autoinmunidad.
A continuación, se explica cada uno de estos conceptos en lenguaje accesible, conservando los números de referencia que aparecen en el estudio original.
Theresa Deisher explica como hay residuos de ADN de lineas celulares de fetos humanos abortados en las vacunas que pueden causar todo tipo de problemas en los vacunados https://www.bitchute.com/embed/1yQc8HbJI9lN
2. Contenido de algunas vacunas
Cuando pensamos en una vacuna, imaginamos un líquido que contiene el virus atenuado y poco más. Sin embargo, la realidad de la fabricación es más compleja.
Los niveles de ADN contaminante en las vacunas contra el sarampión, rubéola, y paperas, la varicela y algunas vacunas contra la hepatitis A disponibles, superan varias veces la recomendación actual de la Organización Mundial de la Salud
El programa de vacunas expone a los niños pequeños a la inserción de fragmentos de ADN fetal durante un período de desarrollo cerebral significativo pudiendo causar autismo, leucemia, cáncer, esquizofrenia y trastorno bipolar y/o posiblemente contribuir al alarmante aumento de disforia de género.
Para producir virus vacunales en cantidades suficientes, los fabricantes necesitan cultivar los virus dentro de células vivas, ya que los virus solo pueden replicarse infectando células. Estas células se mantienen en el laboratorio formando lo que se denomina una línea celular (1). El problema es que cuando se extrae el virus, también se recogen restos de las células fetales que sirvieron para cultivarlo, incluyendo fragmentos de su ADN.
En el caso de varias vacunas administradas rutinariamente a niños —incluyendo la triple vírica SRP (sarampión, paperas y rubéola, conocida como MMR en inglés)— las líneas celulares utilizadas provienen de fetos humanos abortados. Los fabricantes las denominan «líneas celulares diploides humanas», y dos de las más utilizadas se llaman WI-38 y MRC-5, ambas derivadas de tejido pulmonar fetal (1).
Deisher et al., – Epidemiologic and Molecular Relationship Between Vaccine Manufacture and Autism Spectrum Disorder Prevalence – PMID: 26103708 – https://pubmed.ncbi.nlm.nih.gov/26103708/
3. El problema de los residuos de ADN fetal humano
3.1. ¿De dónde viene ese ADN?
Cuando un fabricante cultiva virus en células WI-38 o MRC-5, el producto final —el vial que se inyecta a un niño— contiene inevitablemente fragmentos del ADN de esas células fetales. No es un efecto secundario inesperado: es una consecuencia del proceso de fabricación (1, 6).

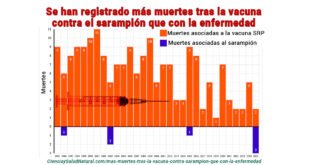
Los efectos secundarios de la vacuna contra el Sarampión, Rubeola y Paperas, SRP (MMR en EE.UU.) incluyen convulsiones, que ocurren en aproximadamente 1 de cada 640 niños vacunados, aproximadamente 5 veces más frecuentemente que las convulsiones por infección de sarampión, sepa como eximir a sus hijos de esta vacuna. Este compendio de estudios de expertos, contiene la suficiente evidencia para que los padres puedan presentar a sus médicos y abogados y prevenir que su hijos sean intoxicados con vacunas que no tienen los suficientes estudios de seguridad como corresponde. Tambien sirve para educar a los médicos sin pensamiento crítico. Descargar libro click aqui
3.2. Cantidades que superan los límites recomendados
La Organización Mundial de la Salud (OMS) ha establecido un límite máximo recomendado de 10 nanogramos (ng) de ADN celular residual por dosis para vacunas producidas en líneas celulares continuas (4). Sin embargo, este límite no se basó en estudios empíricos que demostraran su seguridad, sino que fue el resultado de negociaciones entre reguladores y fabricantes (2, 3, 4). De hecho, el límite se ha ido relajando con el tiempo: de 10 picogramos iniciales (2), pasó a 100 picogramos (3), y finalmente a 10 nanogramos en 1997 (4).
Lo alarmante es que varias vacunas comercializadas en Estados Unidos superan ampliamente ese límite:
- La vacuna monovalente contra la rubéola (disponible en EE.UU. hasta 2011) contenía más de 150 ng de ADN celular por dosis, equivalente al ADN de más de 22.000 células humanas (6, 11).
- Una vacuna contra la hepatitis A fabricada con MRC-5 contiene más de 300 ng por dosis (11).
- La vacuna contra la varicela contiene más de 2 microgramos (2000 ng) de ADN fetal MRC-5 por dosis, según las propias mediciones del fabricante (12).
3.3. Fragmentación: un intento de solución que pudo empeorar las cosas
Conscientes del riesgo de que fragmentos largos de ADN pudieran contener genes cancerígenos completos, las agencias reguladoras recomendaron fragmentar el ADN contaminante en trozos pequeños —idealmente de 200 pares de bases o menos— mediante digestión enzimática, tratamiento químico o radiación gamma (5, 9, 10).
Sin embargo, los autores señalan una ironía peligrosa: la ciencia ha demostrado que los fragmentos cortos de ADN se integran en el genoma con mucha mayor eficiencia que los fragmentos largos. La integración es máxima cuando los fragmentos miden entre 100 y 1.000 pares de bases (13, 14). Es decir, la solución aplicada pudo haber incrementado el peligro en lugar de reducirlo.

Este documento contiene la suficiente evidencia cientifica (más de 150 referencias) para que las madres puedan presentar a sus médicos y abogados y lograr exenciones para prevenir ser dañadas con vacunas o inyecciones génicas, que no tienen los suficientes estudios de seguridad como corresponde. Tambien sirve para educar a los médicos sin pensamiento crítico. descargar libro, click aqui
4. Mutagénesis insercional: Cuando el ADN extraño se introduce en nuestros genes
La mutagénesis insercional es un fenómeno científicamente documentado: fragmentos de ADN externo pueden introducirse en el ADN de las células del receptor, alterando sus genes y provocando mutaciones.
4.1. ¿Cómo entra el ADN contaminante a nuestras células?
Las células de los mamíferos —incluyendo las humanas— pueden captar fragmentos de ADN del entorno mediante un proceso llamado endocitosis mediada por receptores (15). Los estudios experimentales han demostrado que:
- La captación es más eficiente cuando la concentración de ADN externo es baja (15).
- Alcanza su punto máximo aproximadamente 2 horas después de la exposición (16).
- Los fragmentos de ADN pueden alcanzar concentraciones dentro del núcleo celular iguales o superiores a las del medio externo en 2 a 4 horas (17, 21, 23, 24).
- En experimentos con fragmentos de ADN placentario de 500 pares de bases, aproximadamente el 4% del contenido genómico celular se incorporaba por hora de incubación; entre un 40% y un 50% del ADN añadido era captado por las células, y entre un 10% y un 20% llegaba al núcleo (15).
Esto significa que el ADN fetal contaminante inyectado con una vacuna tiene una vía biológicamente plausible para llegar al núcleo de las células del niño y potencialmente integrarse en su genoma.
4.2. La conexión con el autismo y otros trastornos del neurodesarrollo
Los autores presentan un argumento escalonado que conecta los residuos de ADN fetal con los trastornos del espectro autista (TEA):
Primero, se ha descubierto que los genes implicados en la reparación de roturas de doble cadena del ADN (double strand break repair (DSB, por sus siglas en inglés)) están expresados de forma anormal en niños con autismo (25). La reparación de DSB es un proceso normal que ocurre cuando el ADN se rompe, pero si este mecanismo funciona mal, el ADN extraño puede insertarse más fácilmente durante el proceso de reparación.
Segundo, los puntos calientes de recombinación (zonas del genoma donde ocurren con mayor frecuencia las roturas y reparaciones del ADN) están concentrados precisamente en los genes que se han asociado con el autismo (11, 40). Esto crea una susceptibilidad subyacente: cuando un niño con esta característica genética recibe fragmentos de ADN fetal, estos pueden insertarse preferentemente en genes ya vinculados al TEA.
Este documento contiene la suficiente evidencia científica (más de 50) para que las madres puedan presentar a sus médicos y abogados y prevenir sus hijas e hijos sean dañados con vacunas que no tienen los suficientes estudios de seguridad como corresponde. Tambien sirve para educar a los médicos sin pensamiento crítico. No espere hasta último momento para estar protegida… descargar desde: https://cienciaysaludnatural.com/recursos

Tercero, se han identificado más de 350 genes asociados con los trastornos del espectro autista (33, 34, 35). Hasta un 10% de los casos de autismo simplex (sin antecedentes familiares) presentan deleciones o duplicaciones genéticas de novo, es decir, mutaciones no heredadas de los padres (36, 37). Los autores argumentan que esta cifra del 10% probablemente subestima el fenómeno, ya que las técnicas actuales solo detectan variaciones grandes en el número de copias (CNV) y no capturan mutaciones más pequeñas (38).
Cuarto, el mismo patrón de mutaciones de novo se observa en otros trastornos del neurodesarrollo infantil:
- esquizofrenia de inicio temprano (que afecta al 0,4% de la población) (44),
- trastorno bipolar pediátrico (45, 46) y
- discapacidad intelectual (48, 49, 50, 51).
La persistencia de estas enfermedades a pesar de que reducen la capacidad reproductiva sugiere que hay componentes ambientales y genómicos no hereditarios importantes (47).
Quinto, los autores proponen un mecanismo para explicar cómo una mutación en una sola célula podría causar un trastorno cerebral difuso como el autismo. La hematología moderna ha demostrado que nuestro sistema sanguíneo es en gran medida clonal: de los millones de células madre hematopoyéticas que tenemos, solo entre 7 y 8 están activas produciendo los billones de células sanguíneas.
En muchas personas, 1 o 2 células madre producen hasta el 90% de todas las células sanguíneas (88, 89). Si una de estas células madre capta ADN fetal de una vacuna y sufre una mutación, esa mutación se propagará a una enorme proporción de las células sanguíneas. Además, las células gliales del cerebro —fundamentales para la señalización neuronal y la respuesta inmunitaria cerebral— se generan a partir de células madre hematopoyéticas y pueden ser reemplazadas durante la vida. Si estas células gliales de reemplazo portan la mutación, el resultado sería una disfunción cerebral difusa (88, 89).

5. Autoinmunidad: Cuando el cuerpo se ataca a sí mismo
El segundo gran mecanismo de daño identificado por los autores es la autoinmunidad. La lógica es la siguiente:
Cuando se inyecta a un niño fragmentos de ADN fetal humano —que es «no propio», es decir, genéticamente distinto al ADN del niño—, el sistema inmunitario puede generar anticuerpos contra ese ADN extraño. Pero como el ADN fetal tiene grandes regiones de similitud con el ADN del propio niño, esos anticuerpos pueden acabar atacando también los tejidos del niño.
Los autores citan estudios que demuestran que:
- El 62,5% de los niños con autismo presentan anticuerpos antineuronales en sangre, frente a solo el 5% de los niños sanos de control (79).
- La frecuencia de estos anticuerpos es especialmente alta en niñas con autismo: 90% frente al 53,3% en niños varones (79).
- Los niños con autismo presentan anticuerpos contra el ADN humano que los niños no autistas no tienen, lo que sugiere una reacción autoinmune dirigida contra su propio material genético (79, 80, 81, 85, 86, 87).

Respuesta de ANMAT a solicitud de información confirmando el uso de linea de células de fetos humanos abortados en las vacunas del calendartio escolar argentino
El estudio señala que aproximadamente el 40% de los casos de autismo regresivo (aquel en que el niño se desarrolla normalmente y luego pierde habilidades) podrían tener un origen autoinmune, mientras que el 60% restante correspondería a casos de mutagénesis insercional en células madre sanguíneas (88, 89).
Un dato adicional relevante: durante los primeros años de vida, el cerebro humano experimenta una muerte neuronal masiva como parte del desarrollo normal —se eliminan circuitos sobrantes y se podan sinapsis no utilizadas (82, 83)—. Durante este proceso, el ADN de las neuronas que mueren queda expuesto extracelularmente, proporcionando una diana perfecta para los anticuerpos generados tras la exposición al ADN fetal de las vacunas.

6. El papel del retrovirus HERV-K
Algunas vacunas —incluyendo las de varicela y sarampión/paperas/rubéola— no solo contienen ADN fetal humano, sino también fragmentos de un retrovirus endógeno humano llamado HERV-K (62).
¿Qué significa esto? Los retrovirus endógenos son virus antiguos que en algún momento de la evolución se integraron en el genoma humano y se transmiten de generación en generación. Generalmente se consideran inactivos e inofensivos. Sin embargo, la investigación reciente ha demostrado que:
- HERV-K puede reactivarse e incluso mantenerse activo en humanos actuales (63, 64, 65, 66, 67).
- El fragmento de HERV-K presente en las vacunas probablemente codifica la integrasa (la enzima que permite al virus insertarse en el ADN) o la proteína de envoltura (que puede causar neuroinflamación) (64, 77, 78).
- HERV-K activo se integra preferentemente en unidades de transcripción génica y regiones reguladoras, es decir, en zonas del genoma donde puede causar más daño (68).
- Se han encontrado niveles elevados de transcritos de HERV-K en cerebros de pacientes con esquizofrenia y trastorno bipolar (69, 70), así como en células sanguíneas de pacientes con trastornos del espectro autista (71).
- HERV-K pertenece a la misma familia de retrovirus que el virus MMLV (75), utilizado en un ensayo de terapia génica que resultó en leucemia en 4 de 9 niños tratados, como se explica a continuación (76).
7. El momento crítico: La ventana de vulnerabilidad en la infancia
Los autores subrayan un punto cronológico crucial. Con el calendario de vacunación actual, un niño puede recibir hasta 7 o más vacunas contaminadas con ADN fetal antes de cumplir los 2 o 3 años. En los años 90, esa cifra era de solo 2 (6).
Esto coincide con el período más activo del desarrollo cerebral: establecimiento de circuitos neuronales, poda sináptica masiva y muerte neuronal programada a gran escala (82, 83). Es precisamente durante esta ventana de vulnerabilidad cuando la exposición repetida a ADN fetal extraño y a fragmentos retrovirales podría causar el máximo daño, ya sea por inserción mutagénica o por desencadenamiento autoinmune.

En los EE.UU, el autismo ha aumentado en 3 años distintos, llamados puntos de cambio. El primer punto de cambio ocurrió en 1981, el segundo en 1981, y el tercero en 1996. Estos picos coinciden con la introducción de vacunas que se producen en células fetales humanas. En 1979, la célula fetal humana producida MMR II (SRP – tripleviral, contra Sarampión, Rubeola, Paperas). Las campañas de MMR II se elevaron de un 49% para los niños nacidos antes de 1987 a más del 82% para los niños nacidos en 1989 y posteriores. También se introdujo una segunda dosis de MMR II en el calendario de vacunación de los niños nacidos en 1988 y posteriores. El tercer punto de cambio corresponde a la aprobación de Varivax (viruela) producida con células fetales humanas en 1995 (véase la figura a arriba). https://www.soundchoice.org/autism/
https://www.soundchoice.org/wp-content/uploads/2012/08/Regressive_Autism_Prevalence.pdf
8. El ejemplo de la terapia génica que salió mal
Los autores recuerdan un precedente estremecedor. En un ensayo de terapia génica para niños con inmunodeficiencia combinada grave (SCID, también conocida como «enfermedad del niño burbuja»), los expertos de la División de Terapia Génica de la FDA estimaron que el riesgo de que los fragmentos de ADN retroviral y humano causaran mutaciones y cáncer era de 1 entre un billón (76).
La realidad fue devastadoraamente distinta: 4 de los 9 niños tratados (44%) desarrollaron leucemia (76). Los fragmentos de ADN se insertaron en sus genomas y activaron oncogenes. La diferencia entre el riesgo estimado por los reguladores (0,0000000001%) y el riesgo real observado (44%) habla por sí sola sobre la fiabilidad de las evaluaciones oficiales de seguridad.

La seguridad de las vacunas contra la hepatitis B que se administran a los recién nacidos no se ha probado en un solo ensayo clínico controlado aleatorio con placebo inerte como se manifiesta en los propios prospectos y tiene sobredosis de aluminio neurotóxico. Este compendio de estudios de expertos, contiene la suficiente evidencia para que los padres puedan presentar a sus médicos y abogados y prevenir que su hijos sean intoxicados con vacunas que no tienen los suficientes estudios de seguridad como corresponde. Tambien sirve para educar a los médicos sin pensamiento crítico. Descargar libro click aqui
9. Recomendaciones de los autores
El estudio concluye con varias recomendaciones concretas:
- Transparencia en el etiquetado: Que los prospectos de las vacunas incluyan información sobre las cantidades de ADN fetal residual presentes en cada dosis.
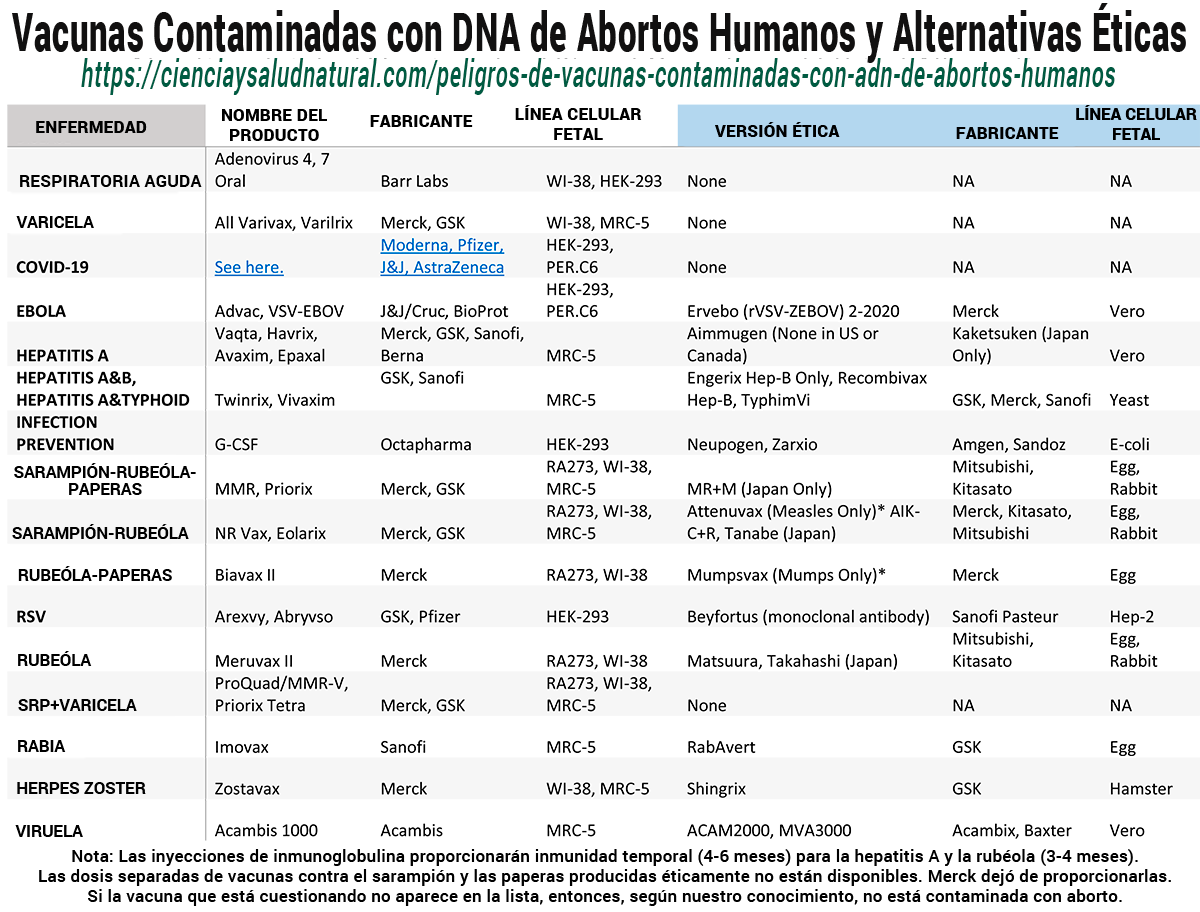
- Alternativas disponibles: Existen vacunas fabricadas en líneas celulares animales, de insectos o de plantas que eliminan el riesgo de contaminación con ADN humano y retrovirus. Por ejemplo, en Japón se utiliza una vacuna contra la rubéola producida en células de riñón de conejo (cepa Takahashi) (84).
- Investigación independiente: Los autores consideran que los peligros de los fragmentos retrovirales y del ADN diploide humano residual constituyen un riesgo no estudiado para los receptores de vacunas, y que el conjunto de la literatura científica demuestra claramente la alta probabilidad de peligros por mutagénesis insercional y/o autoinmunidad. En sus propias palabras, se trata de «un asunto que indudablemente exige una investigación epidemiológica y científica seria» (6).
- Estudios en curso: El Sound Choice Pharmaceutical Institute está llevando a cabo un ensayo clínico observacional para determinar si los niños con autismo presentan respuestas inmunitarias contra los contaminantes fetales específicos presentes en las vacunas (ADN de las líneas MRC-5 y WI-38), así como contra su propio ADN (autoinmunidad), en comparación con niños de desarrollo típico.

Descargar desde https://red.cienciaysaludnatural.com/
10. Referencias
- Gordon K, et al. Immortality, but not oncogenic transformation, of primary human cells leads to epigenetic reprogramming of DNA methylation and gene expression. Nucleic Acids Res. 2014;42:3529-3541.
- Petricciani JC, Horaud FN. DNA, dragons and sanity. Biologicals. 1995;23:233-8.
- WHO Expert Committee on Biological Standardization. WHO Technical Report Series 878. Ginebra: OMS.
- WHO Expert Committee on Biological Standardization. WHO Technical Report Series 941. Ginebra: OMS.
- Hahn WC, et al. Creation of human tumour cells with defined genetic elements. Nature. 1999;400:464-468.
- Deisher T, et al. Epidemiologic and Molecular Relationship Between Vaccine Manufacture and Autism Spectrum Disorder Prevalence. Issues in Law & Medicine. 2015;30(1):47-70.
- Hahn WC, Weinberg RA. Rules for making human tumor cells. N Engl J Med. 2002;347:1593-1603.
- Valastyan S, Weinberg RA. Tumor metastasis: molecular insights and evolving. Cell. 2011;147:275-292.
- Weinberg RA. How cancer arises. Sci Am. 1996;275:62-70.
- Brown F, et al. Safety of Biological Products Prepared from Mammalian Cell Culture. Developments in Biologicals. 1998;93.
- Deisher T, et al. Epidemiologic and Molecular Relationship Between Vaccine Manufacture and Autism Spectrum Disorder Prevalence. Issues in Law & Medicine. 2015;30(1):47-70.
- FDA. Varicella vaccine package insert – residual DNA information. Disponible en: http://www.fda.gov/downloads/biologicsbloodvaccines/vaccines/approvedproducts/ucml42826.pdf
- Lukacs GL, Haggie P, Seksek O, Lechardeur D, Freedman N, Verkman AS. Size-dependent DNA mobility in cytoplasm and nucleus. J Biol Chem. 2000;275(3):1625-9.
- Tsuchiya H, Harashima H, Kamiya H. Factors affecting SFHR gene correction efficiency with single-stranded DNA fragment. Biochem Biophys Res Commun. 2005;336(4):1194-2000.
- Yakubov LA, Deeva EA, Zarytova VF, Ivanova EM, Ryte AS, Yurchenko LY, Vlassov V. Mechanisms of oligonucleotide uptake by cells: Involvement of specific receptors? Proc Natl Acad Sci. 1989;86:6454-6458.
- Vlassov V, Balakireva LA, Yakubov LA. Transport of oligonucleotides across natural and model membranes. Biochim Biophys Acta. 1994;1197(2):95-108.
- Orson FM, Thomas DW, McShan WM, Kessler DJ, Hogan ME. Oligonucleotide inhibition of IL2R alpha mRNA transcription by promoter region collinear triplex formation in lymphocytes. Nucleic Acids Res. 1991;19(12):3435-41.
- Zamecnik PC, Goodchild J, Taguchi Y, Sarin PS. Inhibition of replication and expression of human T-cell lymphotropic virus type III in cultured cells by exogenous synthetic oligonucleotides complementary to viral RNA. Proc Natl Acad Sci USA. 1986;83(12):4143-6.
- Loke SL, Stein CA, Zhang XH, Mori K, Nakanishi M, Subasinghe C, Cohen JS, Neckers LM. Characterization of oligonucleotide transport into living cells. Proc Natl Acad Sci USA. 1989;86(10):3474-8.
- Stein S, Ott MG, Schultze-Strasser S, Jauch A, Burwinkel B, Kinner A, et al. Genomic instability and myelodysplasia with monosomy 7 consequent to EVI1 activation after gene therapy for chronic granulomatous disease. Nat Med. 2010;16(2):198-204.
- Postel EH, Flint SJ, Kessler DJ, Hogan ME. Evidence that a triplex-forming oligodeoxyribonucleotide binds to the c-myc promoter in HeLa cells, thereby reducing c-myc mRNA levels. Proc Natl Acad Sci USA. 1991;88(18):8227-31.
- Leonetti JP, Degols G, Clarenc JP, Mechti N, Lebleu B. Cell delivery and mechanisms of action of antisense oligonucleotides. Prog Nucleic Acid Res Mol Biol. 1993;44:143-66.
- Clarenc JP, Lebleu B, Leonetti JP. Characterization of the nuclear binding sites of oligodeoxyribonucleotides and their analogs. J Biol Chem. 1993;268(8):5600-4.
- Zendegui JG, Vasquez KM, Tinsley JH, Kessler DJ, Hogan ME. In vivo stability and kinetics of absorption and disposition of 3′ phosphopropyl amine oligonucleotides. Nucleic Acids Res. 1992;20(2):307-314.
- Outwin E, Carpenter G, Bi W, Withers MA, Lupski JR, O’Driscoll M. Increased RPA1 gene dosage affects genomic stability potentially contributing to 17p13.3 duplication syndrome. PLoS Genet. 2011;7(8):e1002247.
- Maher RL, Branagan AM, Morrical SW. Coordination of DNA replication and recombination activities in the maintenance of genome stability. J Cell Biochem. 2011;112(10):2672-2682.
- Haber JE. DNA recombination: the replication connection. Trends Biochem Sci. 1999;24(7):271-5.
- Tsai AG, Lieber MR. Mechanisms of chromosomal rearrangement in the human genome. BMC Genomics. 2010;11(S1):S1-9.
- Bradley WEC, et al. Hotspots of Large Rare Deletions in the Human Genome. PLOS ONE. 2010;5(2):1-7.
- Ng SH, Maas SA, Petkov PM, Mills KD, Paigen K. Colocalization of somatic and meiotic double strand breaks near the Myc oncogene on mouse chromosome 15. Genes Chromosomes Cancer. 2009;48(10):925-30.
- Holbeck SL, Strathern JN. A role for REV3 in mutagenesis during double-strand break repair in Saccharomyces cerevisiae. Genetics. 1997;147(3):1017-24.
- Hicks WM, Kim M, Haber JE. Increased mutagenesis and unique mutation signature associated with mitotic gene conversion. Science. 2010;329(5987):82-5.
- Weiss LA, Arking DE; Gene Discovery Project of Johns Hopkins & the Autism Consortium; Daly MJ, Chakravarti A. A genome-wide linkage and association scan reveals novel loci for autism. Nature. 2009;461(7265):802-808.
- Marshall CR, Noor A, Vincent JB, Lionel AC, Feuk L, Skaug J, et al. Structural variation of chromosomes in autism spectrum disorder. Am J Hum Genet. 2008;82(2):477-488.
- Jamain S, Quach H, Betancur C, Rastam M, Colineaux C, Gillberg IC, Soderstrom H, Giros B, Leboyer M, Gillberg C, Bourgeron T. Mutations of the X-linked genes encoding neuroligins NLGN3 and NLGN4 are associated with autism. Nat Genet. 2003;34(1):27-29.
- Abrahams BS, Geschwind DH. Advances in autism genetics: on the threshold of a new neurobiology. Nat Rev Genet. 2008;9:341-355.
- Sebat J, Lakshmi B, Malhotra D, Troge J, Lese-Martin C, Walsh T, et al. Strong association of de novo copy number mutations with autism. Science. 2007;316(5823):445-449.
- Sanders SJ, Ercan-Sencicek AG, Hus V, Luo R, Murtha MT, Moreno-De-Luca D, et al. Multiple Recurrent De Novo CNVs, Including Duplications of the 7q11.23 Williams Syndrome Region, Are Strongly Associated with Autism. Neuron. 2011;70(5):863-885.
- Gilman SR, Iossifov I, Levy D, Ronemus M, Wigler M, Vitkup D. Rare de novo variants associated with autism implicate a large functional network of genes involved in formation and function of synapses. Neuron. 2011;70(5):898-907.
- Michaelson JJ, Shi Y, Gujral M, Zheng H, Malhotra D, Jin X, et al. Whole-genome sequencing in autism identifies hot spots for de novo germline mutation. Cell. 2012;151(7):1431-42.
- FDA. Evolving Scientific and Regulatory Perspectives on Cell Substrates for Vaccine Development. 1999. Disponible en: http://www.fda.gov/downloads/biologicsbloodvaccines/newsevents/workshopsmeetingsconferences/transcriptsminutes/ucm056219.pdf
- McNeer NA, Schleifman EB, Cuthbert A, Brehm M, Jackson A, Cheng C, et al. Systemic delivery of triplex-forming PNA and donor DNA by nanoparticles mediates site-specific genome editing of human hematopoietic cells in vivo. Gene Ther. 2013;20(6):658-69.
- Yakubov LA, Rogachev VA, Lakhacheva AC, Bogachev SS, Sebeleva TE, Shilov AG, et al. Natural Human Gene Correction by Small Extracellular Genomic DNA Fragments. Cell Cycle. 2007;6(18):2293-2301.
- Okkels N, et al. Changes in the diagnosed incidence of early onset schizophrenia over four decades. Acta Psychiatr Scand. 2013;127(1):62-68.
- Leibenluft E, Rich BA. Pediatric bipolar disorder. Annu Rev Clin Psychol. 2008;4:163-87.
- Mitchell PB. Bipolar disorder: the shift to overdiagnosis. Can J Psychiatry. 2012;57(11):659-65.
- Newschaffer CJ. Investigating diagnostic substitution and autism prevalence trends. Pediatrics. 2006;117(4):1436-7.
- Robinson PN. Whole-exome sequencing for finding de novo mutations in sporadic mental retardation. Genome Biol. 2010;11(12):144.
- Awadalla P, Gauthier J, Myers RA. Direct Measure of the De Novo Mutation Rate in Autism and Schizophrenia Cohorts. American Journal of Human Genetics. 2010;87(3):316-324.
- O’Roak BJ, et al. Exome sequencing in sporadic autism spectrum disorders identifies severe de novo mutations. Nat Genet. 2011;43(6):585-9.
- de Ligt J, et al. Diagnostic exome sequencing in persons with severe intellectual disability. N Engl J Med. 2012;367(20):1921-9.
- Iossifov I, et al. De novo gene disruptions in children on the autistic spectrum. Neuron. 2012;74(2):285-99.
- Levy D, et al. Rare de novo and transmitted copy-number variation in autistic spectrum disorders. Neuron. 2011;70(5):886-897.
- Girard SL, et al. Increased exonic de novo mutation rate in individuals with schizophrenia. Nat Genet. 2011;43(9):860-3.
- Xu B, et al. De novo gene mutations highlight patterns of genetic and neural complexity in schizophrenia. Nat Genet. 2012;44(12):1365-9.
- Van Den Bossche MJ, et al. Rare copy number variants in neuropsychiatric disorders: Specific phenotype or not? Am J Med Genet B Neuropsychiatr Genet. 2012;159(7):812-22.
- Gilman SR, et al. Rare de novo variants associated with autism implicate a large functional network of genes involved in formation and function of synapses. Neuron. 2011;70(5):898-907.
- Kong A, et al. Rate of de novo mutations and the importance of father’s age to disease risk. Nature. 2012;488(7412):471-5.
- Luo R, et al. Genome-wide transcriptome profiling reveals the functional impact of rare de novo and recurrent CNVs in autism spectrum disorders. Am J Hum Genet. 2012;91(1):38-55.
- Hamdan FF, et al. Excess of de novo deleterious mutations in genes associated with glutamatergic systems in nonsyndromic intellectual disability. Am J Hum Genet. 2011;88(3):306-316.
- O’Roak BJ, et al. Exome sequencing in sporadic autism spectrum disorders identifies severe de novo mutations. Nat Genet. 2011;43(6):585-589.
- Victoria JG, Wang C, Jones MS, Jaing C, McLoughlin K, Gardner S, Delwart EL. Viral nucleic acids in live-attenuated vaccines: detection of minority variants and an adventitious virus. J Virol. 2010;84(12):6033-6040.
- Lee YN, Bieniasz PD. Reconstitution of an infectious human endogenous retrovirus. PLoS Pathog. 2007;3(1):e10.
- Kitamura Y, Ayukawa T, Ishikawa T, Kanda T, Yoshiike K. Human endogenous retrovirus K10 encodes a functional integrase. J Virol. 1996;70(5):3302-3306.
- Dewannieux M, Harper F, Richaud A, Letzelter C, Ribet D, Pierron G, Heidmann T. Identification of an infectious progenitor for the multiple-copy HERV-K human endogenous retroelements. Genome Res. 2006;16(12):1548-1556.
- Dewannieux M, Ribet D, Heidmann T. Risks linked to endogenous retroviruses for vaccine production: a general overview. Biologicals. 2010;38(3):366-70.
- Belshaw R, Dawson AL, Woolven-Allen J, Redding J, Burt A, Tristem M. Genomewide Screening Reveals High Levels of Insertional Polymorphism in the Human Endogenous Retrovirus Family HERV-K(HML2): Implications for Present-Day Activity. J Virol. 2005;79(19):12507-14.
- Brady T, Lee YN, Ronen K, Malani N, Berry CC, Bieniasz PD, Bushman FD. Integration target site selection by a resurrected human endogenous retrovirus. Genes Dev. 2009;23(5):633-642.
- Frank O, Giehl M, Zheng C, Hehlmann R, Leib-Mosch C, Seifarth W. Human endogenous retrovirus expression profiles in samples from brains of patients with schizophrenia and bipolar disorders. J Virol. 2005;79(17):10890-901.
- Diem O, Schaffner M, Seifarth W, Leib-Mosch C. Influence of antipsychotic drugs on human endogenous retrovirus (HERV) transcription in brain cells. PLoS One. 2012;7(1):e30054.
- Balestrieri E, Arpino C, Matteucci C, Sorrentino R, Pica F, Alessandrelli R, et al. HERVs Expression in Autism Spectrum Disorders. PLoS One. 2012;7(11):e48831.
- Tai AK, O’Reilly EJ, Alroy KA, Simon KC, Munger KL, Huber BT, Ascherio A. Human endogenous retrovirus-K18 Env as a risk factor in multiple sclerosis. Mult Scler. 2008;14(9):1175-80.
- Freimanis G, Hooley P, Ejtehadi HD, Ali HA, Veitch A, Rylance PB, et al. A role for human endogenous retrovirus-K (HML-2) in rheumatoid arthritis: investigating mechanisms of pathogenesis. Clin Exp Immunol. 2010;160(3):340-347.
- Dickerson F, Rubalcaba E, Viscidi R, Yang S, Stallings C, Sullens A, et al. Polymorphisms in human endogenous retrovirus K-18 and risk of type 2 diabetes in individuals with schizophrenia. Schizophr Res. 2008;104(1-3):121-6.
- Mason AL, Xu L, Guo L. Identification of a novel retrovirus associated with primary biliary cirrhosis and autoimmune disorders. Patente US 6,787,303. 2004.
- Hacein-Bey-Abina S, Garrigue A, Wang GP, Soulier J, Lim A, Morillon E, et al. Insertional oncogenesis in 4 patients after retrovirus-mediated gene therapy of SCID-X1. J Clin Invest. 2008;118(9):3132-42.
- Antony JM, Van Marle G, Opii W, Butterfield DA, Mallet F, Yong VW, Wallace JL, Deacon RM, Warren K, Power C. Human endogenous retrovirus glycoprotein-mediated induction of redox reactants causes oligodendrocyte death and demyelination. Nat Neurosci. 2004;7:1088-1095.
- Karlsson H, Bachmann S, Schroder J, McArthur J, Torrey EF, Yolken RH. Retroviral RNA identified in the cerebrospinal fluids and brains of individuals with schizophrenia. Proc Natl Acad Sci USA. 2001;98:4634-4639.
- Mostafa G, Al-Ayadhi L. The relationship between the increased frequency of serum antineuronal antibodies and the severity of autism in children. Eur J Paediatr Neurol. 2012;16:

DMSO, Dimetíl Sulfóxido, usos: Accidentes cerebrovasculares, hemorragias cerebrales, lesiones cerebrales y de la columna, parálisis, ataques cardíacos, demencia, amiloidosis, más, descargar desde https://red.cienciaysaludnatural.com/


Parásitos, tratamientos naturales, hierbas y alimentos que pueden librarnos de varios tipos de parásitos: Cáscara de nuez negra, Ajenjo, Clavo de olor y otras, desontoxicación durante y post-tratamiento, como restaurar el microbioma. Más de 200 referencias científicas. Descargar aqui


Zeolita, beneficios para la salud
Restablecer la flora intestinal y proteger la mucosa, recuperación del equilibrio digestivo tras una desparasitación. Reducción en los niveles de estrés oxidativo e inflamación. Reduce la presencia de metales pro-oxidantes (como hierro libre, plomo, mercurio y cadmio). Reduce la neuroinflamación mejorando la función cognitiva. Protege las mitocondrias neuronales, que son particularmente vulnerables a la toxicidad por mercurio o aluminio. DESCARGAR desde: https://red.cienciaysaludnatural.com/page/2/